T-лимфоциты
| Т-лимфоцит | |
|---|---|
 | |
| Ткань | соединительная |
| История дифференцировки клетки | Гемопоэтические стволовые клетки → претимоциты → тимоциты |
| Возможности для дальнейшей дифференцировки | Т-клетки памяти |
T-лимфоциты, или Т-клетки (от лат. thymus «тимус») — лимфоциты, развивающиеся у млекопитающих в тимусе из предшественников — претимоцитов, поступающих в него из красного костного мозга. В тимусе T-лимфоциты дифференцируются, приобретая Т-клеточные рецепторы (ТКР, англ. TCR) и различные корецепторы (поверхностные маркеры). Играют важную роль в приобретённом иммунном ответе. Обеспечивают распознавание и уничтожение клеток, несущих чужеродные антигены, усиливают действие моноцитов, NK-клеток, а также принимают участие в переключении изотипов иммуноглобулинов (в начале иммунного ответа B-клетки синтезируют IgM, позже переключаются на продукцию IgG, IgE, IgA).
Содержание
Типы Т-лимфоцитов
Т-клеточные рецепторы являются основными поверхностными белковыми комплексами Т-лимфоцитов, ответственными за распознавание процессированных антигенов, связанных с молекулами главного комплекса гистосовместимости (ГКГ, англ. Major Histocompatibility Complex (MHC)) на поверхности антигенпрезентирующих клеток. Т-клеточный рецептор связан с другим полипептидным мембранным комплексом, CD3. В функции комплекса CD3 входит передача сигналов в клетку, а также стабилизация Т-клеточного рецептора на поверхности мембраны. Т-клеточный рецептор может ассоциироваться с другими поверхностными белками, его корецепторами. В зависимости от корецептора и выполняемых функций различают два основных типа Т-клеток.
Т-хелперы

Т-хелперы (от англ. helper — помощник) — Т-лимфоциты, главной функцией которых является усиление адаптивного иммунного ответа. Активируют Т-киллеры, B-лимфоциты, моноциты, NK-клетки при прямом контакте, а также гуморально, выделяя цитокины. Основным признаком Т-хелперов служит наличие на поверхности клетки молекулы корецептора CD4. Т-хелперы распознают антигены при взаимодействии их Т-клеточного рецептора с антигеном, связанным с молекулами главного комплекса гистосовместимости II класса (англ. Major Histocompatibility Complex II (MHC-II)).
Т-киллеры
Т-киллеры, цитотоксические T-лимфоциты, CTL (от англ. killer «убийца») — Т-лимфоциты, главной функцией которых является уничтожение повреждённых клеток собственного организма. Мишени Т-киллеров — это клетки, поражённые внутриклеточными паразитами (к которым относятся вирусы и некоторые виды бактерий), опухолевые клетки. Т-киллеры являются главным компонентом антивирусного иммунитета. Основным признаком Т-киллеров служит наличие на поверхности клетки молекулы корецептора CD8. Т-киллеры распознают антигены при взаимодействии их Т-клеточного рецептора с антигеном, связанным с молекулами главного комплекса гистосовместимости I класса (англ. Major Histocompatibility Complex I (MHC-I)).
Т-хелперы и Т-киллеры образуют группу эффекторных Т-лимфоцитов, непосредственно ответственных за иммунный ответ. В то же время существует другая группа клеток, регуляторные Т-лимфоциты, функция которых заключается в регулировании активности эффекторных Т-лимфоцитов. Модулируя силу и продолжительность иммунного ответа через регуляцию активности Т-эффекторных клеток, регуляторные Т-клетки поддерживают толерантность к собственным антигенам организма и предотвращают развитие аутоиммунных заболеваний. Существуют несколько механизмов супрессии: прямой, при непосредственном контакте между клетками, и дистантный, осуществляющийся на расстоянии — например, через растворимые цитокины.
γδ Т-лимфоциты
Т-лимфоциты представляют собой небольшую популяцию клеток с видоизменённым Т-клеточным рецептором. В отличие от большинства других Т-клеток, рецептор которых образован и субъединицами, Т-клеточный рецептор -лимфоцитов образован и субъединицами. Данные субъединицы не взаимодействуют с пептидными антигенами, презентированными белками ГКГ. Предполагается, что Т-лимфоциты участвуют в узнавании липидных антигенов.





Т-супрессоры
Т-лимфоциты, обеспечивающие центральную регуляцию иммунного ответа.
Дифференциация в тимусе
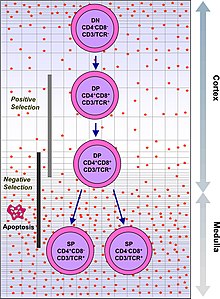
Все Т-клетки берут своё начало от гемопоэтических стволовых клеток красного костного мозга, которые мигрируют в тимус и дифференциируются в незрелые тимоциты. Тимус создаёт микросреду, необходимую для развития полностью функционального репертуара Т-клеток, который является ГКГ-ограниченным и толерантным к самому себе.
Дифференциация тимоцитов разделяется на разные стадии в зависимости от экспрессии различных поверхностных маркеров (антигенов). На самой ранней стадии тимоциты не экспрессируют корецепторы CD4 и CD8 и поэтому классифицируются как двойные негативные (англ. Double Negative (DN)) (CD4-CD8-). На следующей стадии тимоциты экспрессируют оба корецептора и называются двойными позитивными (англ. Double Positive (DP)) (СD4+CD8+). Наконец на финальной стадии происходит селекция клеток, которые экспрессируют только один из корецепторов (англ. Single Positive (SP)): или (CD4+), или (CD8+).
Раннюю стадию можно разделить на несколько подстадий. Так, на подстадии DN1 (англ. Double Negative 1) тимоциты имеют следующую комбинацию маркеров: CD44+CD25-CD117+. Клетки с данной комбинацией маркеров ещё называют ранними лимфоидными предшественниками (англ. Early Lymphoid Progenitors (ELP)). Прогрессируя в своей дифференциации, ELP активно делятся и окончательно теряют способность трансформироваться в другие типы клеток (например В-лимфоциты или миелоидные клетки). Переходя на подстадию DN2 (англ. Double Negative 2), тимоциты экспрессируют CD44+CD25+CD117+ и становятся ранними Т-клеточными предшественниками (англ. Early T-cell Progenitors (ETP)). В течение DN3 подстадии (англ. Double Negative 3) ETP клетки имеют комбинацию CD44-CD25+ и вступают в процесс β-селекции.
β-селекция
Гены Т-клеточного рецептора состоят из повторяющихся сегментов, принадлежащих к трём классам: V (англ. variable), D (англ. diversity) и J (англ. joining). В процессе соматической рекомбинации генные сегменты, по одному из каждого класса, соединяются вместе (V(D)J-рекомбинация). Случайное объединение последовательностей сегментов V(D)J приводит к появлению уникальных последовательностей вариабельных доменов каждой из цепей рецептора. Случайный характер образования последовательностей вариабельных доменов позволяет генерировать Т-клетки, способные распознавать большое количество различных антигенов, и, как следствие, обеспечивать более эффективную защиту против быстро эволюционирующих патогенов. Однако этот же механизм зачастую приводит к образованию нефункциональных субъединиц Т-клеточного рецептора. Гены, кодирующие β-субъединицу рецептора, первыми подвергаются рекомбинации в DN3-клетках. Чтобы исключить возможность образования нефункционального пептида, β-субъединица образует комплекс с инвариабельной α-субъединицей пре-T-клеточного рецептора, формируя т. н. пре-T-клеточный рецептор (пре-ТКР). Клетки, неспособные образовывать функциональный пре-ТКР, погибают в результате апоптоза. Тимоциты, успешно прошедшие β-селекцию, переходят на подстадию DN4 (CD44-CD25-) и подвергаются процессу позитивной селекции.
Позитивная селекция
Клетки, экспрессирующие на своей поверхности пре-ТКР все ещё не являются иммунокомпетентными, так как не способны связываться с молекулами главного комплекса гистосовместимости. Для узнавания молекул ГКГ T-клеточным рецептором необходимо наличие корецепторов CD4 и CD8 на поверхности тимоцитов. Образование комплекса между пре-ТКР и корецептором CD3 приводит к ингибированию перестроек генов β-субъединицы и в то же время вызывает активацию экспрессии генов CD4 и CD8. Таким образом тимоциты становятся двойными позитивными (DP) (CD4+CD8+). DP-тимоциты активно мигрируют в корковое вещество тимуса, где происходит их взаимодействие с клетками кортикального эпителия, экспрессирующими белки обоих классов ГКГ (MHC-I и MHC-II). Клетки, неспособные взаимодействовать с белками ГКГ кортикального эпителия, подвергаются апоптозу, в то время как клетки, успешно осуществившие такое взаимодействие, начинают активно делиться.
Негативная селекция
Тимоциты, прошедшие позитивную селекцию, начинают мигрировать к кортико-медуллярной границе тимуса. Попадая в медуллу, тимоциты взаимодействуют с собственными антигенами организма, презентированными в комплексе с белками ГКГ на медуллярных тимических эпителиальных клетках (мТЭК). Тимоциты, активно взаимодействующие с собственными антигенами, подвергаются апоптозу. Негативная селекция предотвращает появление самоактивирующихся Т-клеток, способных вызывать аутоиммунные заболевания, являясь важным элементом иммунологической толерантности организма.
Активация

Т-лимфоциты, успешно прошедшие позитивную и негативную селекцию в тимусе, попавшие на периферию организма, но не имевшие контакта с антигеном, называются наивными Т-клетками (англ. Naive T cells). Основной функцией наивных Т-клеток является реакция на патогены, прежде не известные иммунной системе организма. После того как наивные Т-клетки распознают антиген, они становятся активированными. Активированные клетки начинают активно делиться, образуя клон. Некоторые из клеток этого клона превращаются в эффекторные Т-клетки, которые выполняют функции, специфичные для данного типа лимфоцита (например, выделяют цитокины в случае Т-хелперов или же лизируют поражённые клетки в случае Т-киллеров). Другая часть активированных клеток трансформируется в Т-клетки памяти. Клетки памяти сохраняются в неактивной форме после первичного контакта с антигеном до тех пор, пока не наступает повторное взаимодействие с тем же антигеном. Таким образом, Т-клетки памяти хранят информацию о ранее действовавших антигенах и обеспечивают вторичный иммунный ответ, осуществляющийся в более короткие сроки, чем первичный.
Взаимодействие Т-клеточного рецептора и корецепторов (СD4, CD8) с главным комплексом гистосовместимости важно для успешной активации наивных Т-клеток, однако его самого по себе недостаточно для дифференциации в эффекторные клетки. Для последующей пролиферации активированных клеток необходимо взаимодействие т. н. костимулирующих молекул. Для Т-хелперов такими молекулами являются рецептор CD28 на поверхности Т-клетки и иммуноглобулин B7 на поверхности антигенпрезентирующей клетки.